
В Корее нашли натуральное вещество, подавляющий рост раковых клеток
- 21 час назад
- 6 мин. чтения
Корреспондент Гу Бон Хёк
- Впервые в мире раскрыт механизм подавления «mTOR» — ключевого белка, ответственного за рост рака
- Метаболит «13-HODE» подавляет активность белка mTOR, сдерживая рост раковых клеток

Обложка майского номера международного научного журнала «Cell Chemical Biology», в котором опубликованы результаты данного исследования [Предоставлено KAIST]
Белок mTOR в организме человека чрезмерно активируется в раковых клетках, вызывая пролиферацию опухолей и метастаз. Однако механизм действия молекул, регулирующих активность белка mTOR, до сих пор не был выяснен.
2 мая KAIST сообщил, что исследовательская группа под руководством профессора Ким Се Юна из факультета биологических наук в сотрудничестве с командой во главе с профессором Бён Ён Чжу из факультета фармакологии Университета Корё обнаружила, что 13-HODE (продукт метаболизма жирных кислот), являющийся метаболитом липидов в организме, подавляет активность mTOR, ключевого регулятора роста раковых клеток.
Данное исследование открывает возможности для разработки стратегий противоракового лечения нового поколения.
Исследовательская группа сосредоточила внимание на веществах, способных связываться с белком mTOR, в частности на природных метаболитах, самостоятельно вырабатываемых нашим организмом. В результате обширного метаболического скрининга (технологии массового анализа метаболитов в организме) было обнаружено, что «13-HODE» — липидный метаболит, образующийся в результате трансформации жиров в организме — непосредственно присоединяется к активному участку белка mTOR и блокирует его функционирование в раковых клетках.
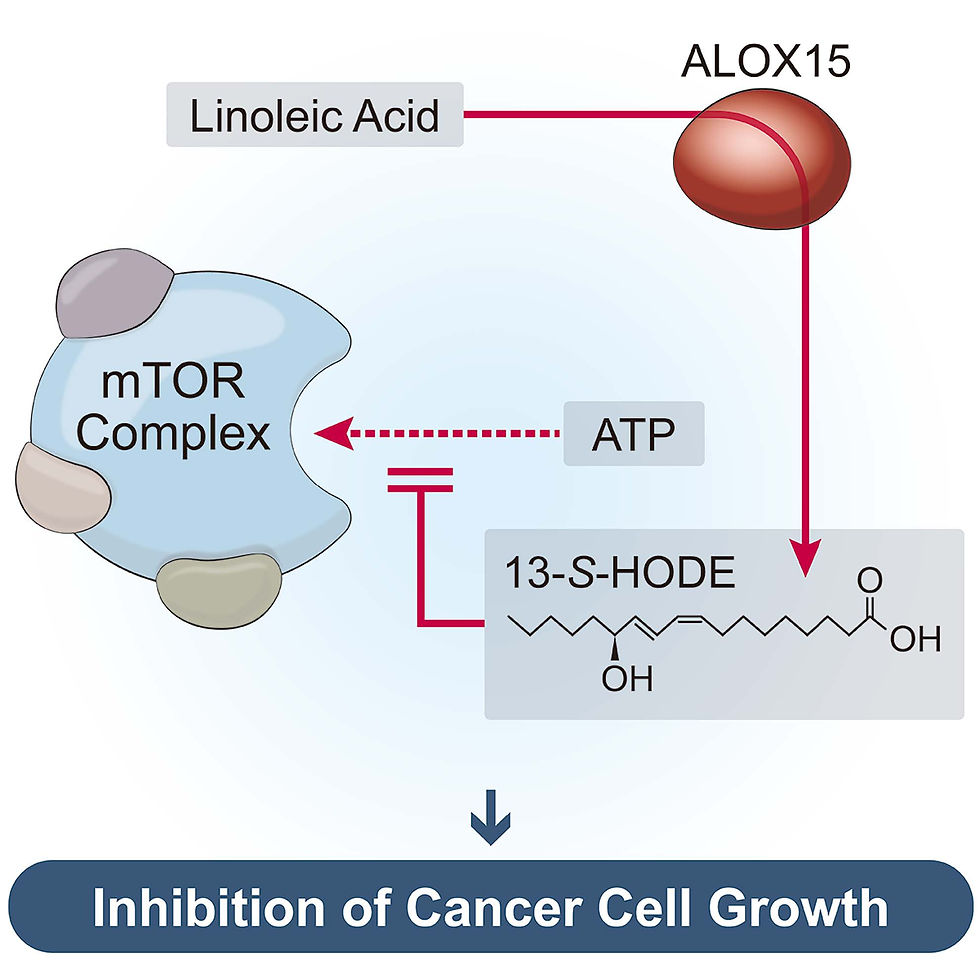
Молекула 13-HODE образуется в нашем организме в процессе метаболизма линолевой кислоты - ненасыщенной жирной кислоты, незаменимые для человеческого организма - которая в изобилии содержится в растительных маслах и т. п. В ходе этого процесса фермент «ALOX15» (фермент, индуцирующий окисление жирных кислот) окисляет линолевую кислоту, образуя 13-HODE.
Аннотация в графиках. Краткое содержание: Рост клеток регулируется сигнальным путем mTOR, однако остается неясным, каким образом метаболизм непосредственно влияет на этот путь. Парк, Ким и др. выявили, что метаболит линолевой кислоты 13-S-HODE является эндогенным ингибитором mTOR, что связывает метаболизм полиненасыщенных жирных кислот с подавлением роста раковых клеток и опухолей.

Рисунок 1. Идентификация 13-S-HODE как метаболита, связанного с mTOR
(A) Схема эксперимента по скринингу эндогенных малых молекул, связывающихся с mTOR. Метаболиты, специфически связывающиеся с иммунопреципитатами Flag-mTOR по сравнению с контролем, были идентифицированы с помощью LC-MS. (B) Хроматограммы экстрагированных ионов FLAG-mTOR (вверху) и отрицательного контроля (посередине) на 6,51 мин в отрицательном режиме. Обильный метаболит ([M-H]-:295,2264) в FLAG-mTOR был идентифицирован как 13-S-HODE путем сравнения с точными определениями молекулярной массы и времени удержания чистого 13-S-HODE ([M-H]-: 295,2271, внизу) при тех же условиях LC-MS. (C) Химическая структура 13-S-HODE. (D) Связывание меченного биотином 13-S-HODE с эндогенным mTOR, raptor или rictor в лизатах клеток HEK293T, оцененное с помощью пулдауна на стрептавидиновых шариках и иммуноблоттинга. Блоты являются репрезентативными для трех независимых экспериментов.

Рисунок 2. 13-S-HODE непосредственно ингибирует киназную активность mTOR
(A и B) Анализы киназной активности mTOR «ин витро» проводились в отсутствии или в присутствии 13-S-HODE. FLAG-raptor или FLAG-rictor котрансфицировали в клетки HEK293T вместе с mTOR, меченным myc. Киназная активность mTORC1 оценивалась с помощью иммунопреципитатов FLAG-raptor посредством иммуноблоттинга фосфорилирования T389 S6K1 (A). Киназная активность mTORC2 анализировалась с помощью иммунопреципитатов FLAG-rictor на основе фосфорилирования S473 Akt (B). (C) Анализы киназной активности in vitro проводили с использованием рекомбинантного C-концевого участка человеческого mTOR (mTOR1362-2549). Киназную активность mTOR оценивали на основе уровня фосфорилирования S6K1 в положении T389. (D) Связывание меченного биотином 13-S-HODE с рекомбинантным mTOR1362-2549 оценивали с помощью пулдауна на стрептавидиновых шариках и иммуноблоттинга. (E) Анализы киназной активности in vitro проводились с использованием рекомбинантного C-концевого участка человеческого mTOR (mTOR1362-2549) в отсутствии или в присутствии либо линолевой кислоты, либо 13-S-HODE. Все изображения демонстрируют типичные эксперименты, которые были повторены независимо три раза.

Рисунок 3. 13-S-HODE действует как ингибитор, конкурирующий с АТФ
(A) Пептиды, демонстрирующие значительное отклонение, были нанесены на структуру mTOR (PDB: 4JSV) в виде поверхностного представления. Области со значительным снижением поглощения дейтерия в присутствии 13-S-HODE окрашены в зеленый цвет. Ионы магния представлены в виде черных сфер, а АДФ обозначен синим цветом для выделения каталитического центра. (B) Оптимальная поза стыковки 13-S-HODE в каталитическом сайте киназы mTOR. Были обнаружены водородные связи между 13-S-HODE и mTOR (OH-группа 13-S-HODE с Lys2197 и Glu2190, COOH-группа с His2340, Ser2342 и His2247). Цифры обозначают расстояние межмолекулярной водородной связи в А˚. (C и D) Термическая денатурация рекомбинантного домена киназы mTOR измерялась в отсутствии и в присутствии различных концентраций 13-S-HODE, как указано. Показаны типичные кривые производной (dF/dT) для необработанного образца и образца, обработанного различными концентрациями лиганда (C). Средние температуры перехода разворачивания белка (Tm) представлены в виде столбцов (D). Данные представляют собой среднее значение ± стандартное отклонение по результатам не менее трех независимых измерений (*p < 0,05). (E) Для оценки термической стабильности mTOR, связанного с 13-S-HODE, был проведен анализ CETSA. Клетки HEK293T трансфицировали FLAG-mTORWT или FLAG-mTORK 2187A/E2190A. Клеточные лизаты инкубировали с 13-S-HODE в течение 1 часа при 4 °C. В качестве нецелевого белка использовали актин. (F) Активность mTORC1 in vitro иммунопреципитатов raptor, полученных из клеток HEK293, анализировали в присутствии 75 мкМ 13-S-HODE и возрастающих концентраций АТФ. Фосфорилирование S6K1 в положении T389 измеряли с помощью иммуноблоттинга. Показаны типичные эксперименты, которые повторяли независимо три раза (E и F), и все давали схожие результаты.
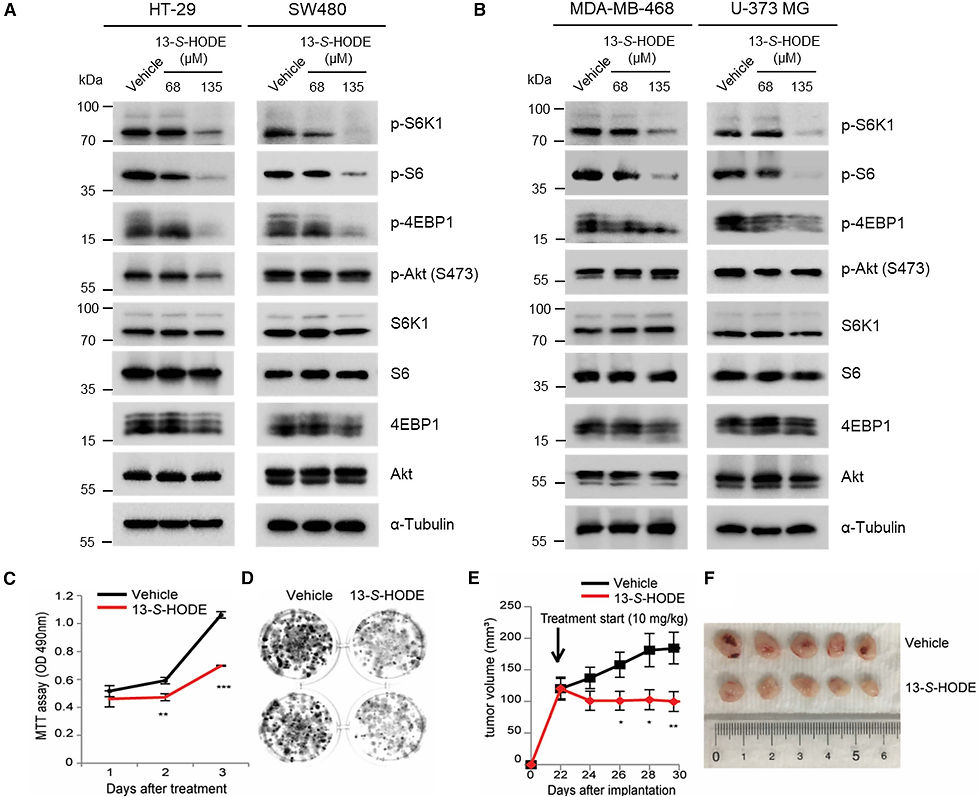
Рисунок 4. 13-S-HODE ингибирует mTOR-сигнализацию и рост раковых клеток
(A) Клетки колоректального рака HT-29 и SW480 обрабатывали DMSO-носителем или 13-S-HODE в течение 48 ч. Клеточные лизаты анализировали с помощью иммуноблоттинга на уровни указанных белков и состояния фосфорилирования. (B) Клетки рака молочной железы MDA-MB-468 и клетки глиобластомы U-373 MG обрабатывали растворителем DMSO или 13-S-HODE в течение 48 ч. Клеточные лизаты анализировали с помощью иммуноблоттинга на содержание указанных белков и степень их фосфорилирования. (C и D) Измеряли жизнеспособность клеток (C) и образование колоний (D) клеток MDA-MB-468, обработанных растворителем DMSO или 135 мкМ 13-S-HODE. Данные представлены в виде среднего значения ± SEM (*p < 0,05; p < 0,01; *p < 0,001, t-критерий Стьюдента). (E и F) Клетки MDA-MB-468 были инокулированы в боковые области голого мыша (n = 6 в группе), и объемы опухолей измерялись в течение 30 дней после инъекции. Растворитель DMSO или 13-S-HODE (10 мг/кг) вводили внутриопухолево пять раз в ксенотрансплантаты каждые 2 дня (E). Данные представлены в виде среднего значения ± стандартная ошибка (*p < 0,05, p < 0,01, *p < 0,001, t-критерий Стьюдента). Типичные изображения ксенотрансплантированных опухолей в день умерщвления (F).
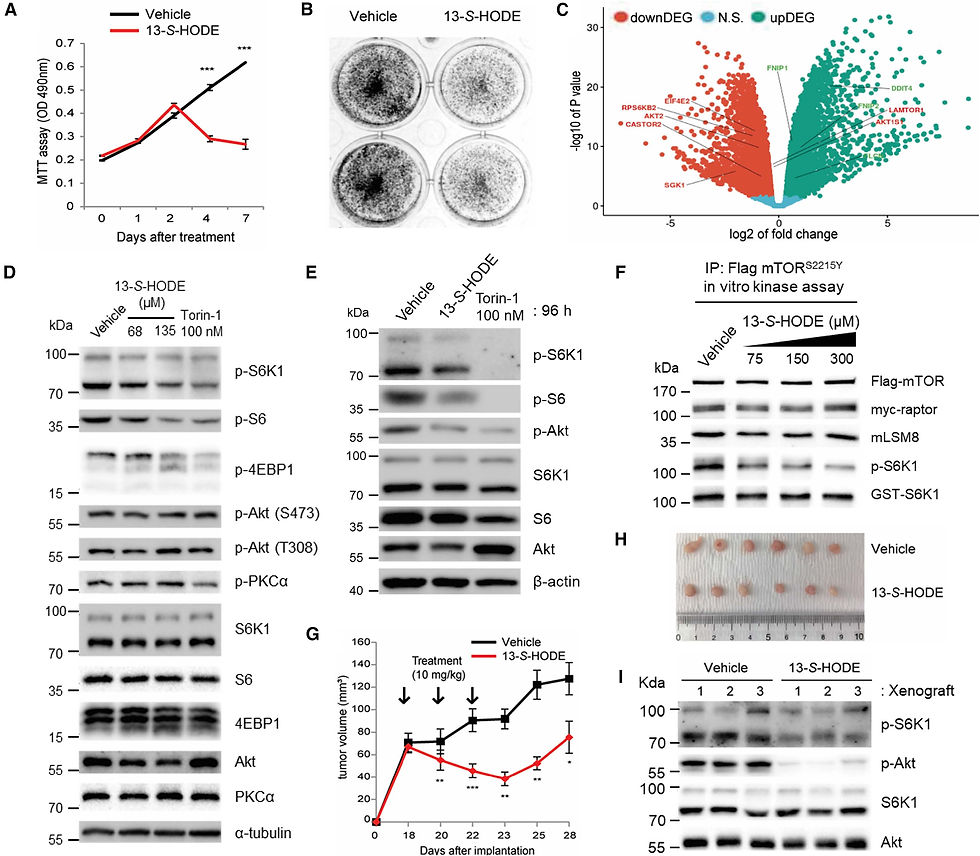
Рисунок 5. 13-S-HODE ингибирует сигнальный путь mTOR и подавляет рост раковых клеток с гиперактивной мутацией mTOR
(A и B) Были измерены жизнеспособность клеток (A) и образование колоний (B) клеток рака эндометрия JHUEM7, обработанных растворителем DMSO или 135 мкМ 13-S-HODE. (C) График «вулкан», показывающий значительные изменения экспрессии генов в ответ на обработку 135 мкМ 13-S-HODE в клетках JHUEM7. (D) Клетки JHUEM7 обрабатывали DMSO-носителем или 13-S-HODE в течение 48 ч. Клеточные лизаты анализировали с помощью иммуноблоттинга на содержание указанных белков и степень их фосфорилирования. (E) Клетки JHUEM7 обрабатывали DMSO-носителем или 135 мкМ 13-S-HODE в течение 96 ч. Клеточные лизаты анализировали методом иммуноблоттинга на содержание указанных белков и степень их фосфорилирования. (F) Активность mTORC1 in vitro иммунопреципитатов mTOR, полученных из клеток HEK293T, экспрессирующих mTORS2215Y, определяли в присутствии 13-S-HODE. (G–I) Раковые клетки JHUEM7 были инокулированы в боковые области обнаженных мышей (n = 6 в группе), и объемы опухолей измерялись в течение 28 дней после инъекции. Носитель DMSO или 13-S-HODE (10 мг/кг) вводили внутриопухолево три раза в ксенотрансплантаты каждые 2 дня (G). Данные представлены в виде среднего значения ± SE (*p < 0,05, p < 0,01, *p < 0,001, t-критерий Стьюдента). Типичные изображения ксенотрансплантатов в день умерщвления (H). Иммуноблот-анализ опухолевых тканей, полученных из ксенотрансплантатов (I). Данные являются типичными для трех независимых экспериментов.
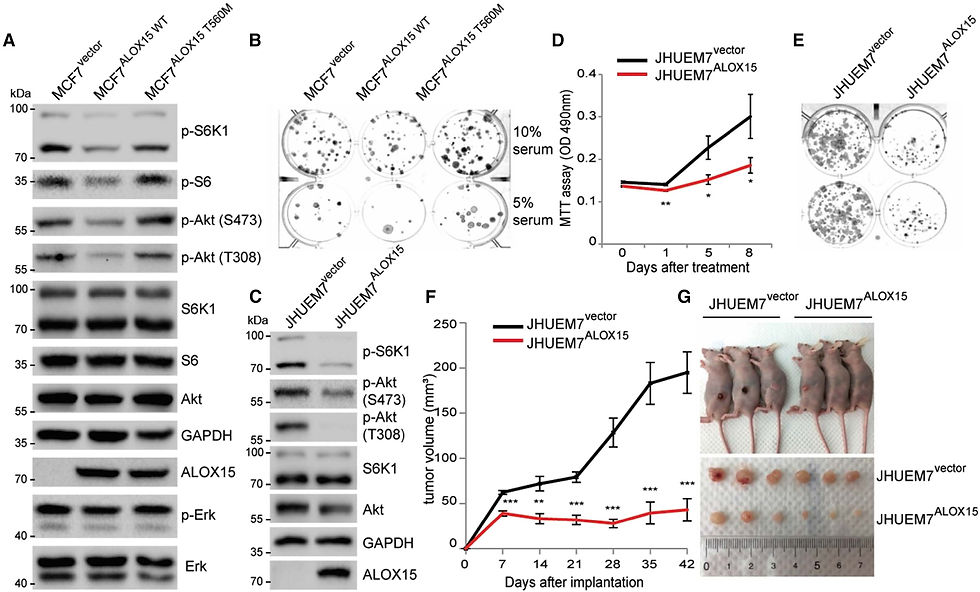
Рисунок 6. Сверхэкспрессия ALOX15 подавляет сигнальный путь mTOR и рост раковых клеток
(A и B) Были созданы клетки MCF7 со стабильной сверхэкспрессией пустого вектора, ALOX15-WT или ALOX15-T560M. Клеточные лизаты анализировали с помощью иммуноблоттинга на уровни указанных белков и состояния фосфорилирования (A). Было измерено образование колоний клеток MCF7 в (A) (B). (C–G) Были получены клетки JHUEM7 со стабильной сверхэкспрессией пустого вектора или ALOX15-WT. Клеточные лизаты анализировали методом иммуноблоттинга на содержание указанных белков и степень их фосфорилирования (C). Были измерены жизнеспособность клеток (D) и образование колоний (E) клеток JHEUM7. Клетки JHUEM7 были инокулированы в боковые области голого мыша (n = 6 в группе), и объемы опухолей измерялись в течение 42 дней после инъекции (F). Данные представлены в виде среднего значения ± стандартная ошибка (*p < 0,05, p < 0,01, *p < 0,001, t-критерий Стьюдента). Типичные изображения опухолей-ксенотрансплантатов в день умерщвления (G).

Профессор Ким Се Юн (слева) из KAIST и профессор Бён Ён Чжэ из Университета Корё [Предоставлено KAIST]
Исследовательская группа доказала, что увеличение выработки ALOX15 и 13-HODE приводит к снижению активности mTOR и подавлению роста раковых клеток.
Суть данного исследования заключается в том, что было выявлено молекулярный механизм (принцип действия в организме), при котором 13-HODE напрямую связывается с mTOR — ключевым белком роста рака — и блокирует его активность.
«Мы считаем, что результаты данного исследования могут быть использованы в качестве базовой технологии для разработки метаболических стратегий лечения, подавляющих рост раковых клеток» - сказал профессор Ким Се Юн, добавив: «В дальнейшем эту концепцию можно будет применять к различным заболеваниям, при которых происходит гиперактивация сигнального пути mTOR, таким как воспаление и старение».
Результаты данного исследования опубликованы в майском номере международного научного журнала по химической биологии «Cell Chemical Biology».



Комментарии